Ptaki są organizmami stałocieplnymi, co jest uwarunkowane odpowiednim przebiegiem procesów metabolicznych. Temperatura ciała ptaków, rozumiana jako odczuwalna temperatura powierzchni ciała, wynosi około 41,5°C (średnio) i może wahać się przedziale plus/minus dwóch stopni (np. temperatury narządów wewnętrznych jak wątroby, odcinków układu pokarmowego czy ośrodkowego układu nerwowego mogą być wyższe), temperatury kończyn – niższe.
Całokształt procesów fizjologicznych, celem których jest utrzymanie homeostazy termicznej ustroju, to między innymi liczne reakcje neuro-hormonalne, procesy endo-enzymatyczne, regulacje szybkości przepływu krwi przez naczynia wewnątrz-narządowe i wokół-narządowe oraz sieć naczyniową w skórze, tak aby średnia wartość termogenenzy (TA) organizmu była stała i w zależności od potrzeb, a więc wpływu środowiska zewnętrznego, regulowana byłaby poprzez zwiększone oddawanie ciepła (gdy jest krytycznie gorąco – zapobieganie hipertermii – HS „heat stress” – stres cieplny) lub jego „kumulowanie” wokół strategicznych narządów gdy jest krytycznie chłodno (zapobieganie hipotermii) (Wertelecki, 2004; Króliczewska i wsp. 2013).
Organizm ptaków bilans termiczny może prowadzić w oparciu o cztery kluczowe procesy, czyli różne formy kontaktu ze środowiskiem zewnętrznym otaczającym go: parowanie, promieniowanie, przewodzenie i konwekcję. Najważniejszym z procesów regulacji bilansu termicznego drobiu jest parowanie, choć ptaki nie mają skórnych gruczołów potowych, największe odparowywanie wody odbywa się u nich z powierzchni błon śluzowych, a w szczególności z powierzchni układu oddechowego. Czynnikiem ochładzającym powierzchnie błon śluzowych jest – woda, która to jest w ten sposób tracona z organizmu. Przyjmuje się, iż organizm zwierząt stałocieplnych w zależności od gatunku, wieku, fazy rozwojowej i fizjologicznej, aż w 68-72% stanowi właśnie woda.

Woda jest też podstawowym prostym związkiem biochemicznym dla budowy wszystkich tkanek organizmu, a w szczególności krwi – tkanki łącznej płynnej. To środowisko wodne organizmu jest sferą, gdzie rozpuszczone są wszystkie składniki organiczne i nieorganiczne, są one w tym środowisku transportowane i gromadzone (rezerwuar). W grupie wspomnianych składników warunkujących przebieg ponad 99% procesów metabolicznych, a w szczególności transportu do i z komórek przez błony komórkowe (a więc i zdolności regulacji osmozy w procesie termogenenzy ustrojowej – stałocieplności) wyróżnia się elektrolity.


Elektrolity z chemicznego punktu widzenia są „monowalentnymi jonami”, kationami jak sód (Na+), potas (K+) oraz anionami jak chlor (Cl–) o masach atomowych wynoszących odpowiednio: 22,9898; 39,102 i 35,453 i stałym ładunku +1, +1 i –1 (Hooge, 2003). Natomiast z punktu widzenia fizycznego są między innymi pierwiastkami łatwo przechodzącymi do roztworów przy jednoczesnym zachowaniu swoich jonowych właściwości. W opinii wielu badaczy zajmujących się tematyką elektrolitów w metabolizmie oprócz ww. trzech podstawowych i kluczowych jonów również w tej grupie znaleźć się mogą niektóre jony dwuwartościowe jak wapniowe, fosforowe, magnezowe, siarczanowe oraz specyficzne jony węglanów i dwuwęglanów (Jamroz, 2001; Hooge, 2003; McNaughton i wsp., 2003).
Elektrolity występują w organizmie ptaków głównie w formie zjonizowanej, nierównomiernie rozdzielone pomiędzy płyny zewnątrzkomórkowe oraz wewnątrzkomórkowe. W medium jakim jest woda ustrojowa stanowiącym dominujący składnik w płynach otaczających komórki (zewnątrzkomórkowych) przede wszystkim identyfikuje się jony sodu (aż 93% jonów płynu), jony chloru oraz wapnia i jony węglanowe, a w płynie wypełniającym komórki, obecny jest potas, magnez, a także jony siarczanowe i fosforanowe (Hooge, 2003).
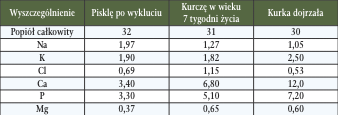
W ciele ptaków orientacyjna koncentracja podstawowych jonów jest w miarę równa w odniesieniu do 1 kg masy ciała, natomiast zawartość wapnia, fosforu czy też magnezu (w formie jonowej) wzrasta w miarę proporcjonalnie do wieku ptaków (Tab. 1) (za Larbier i wsp., 1995).
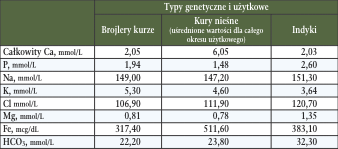
Natomiast przy podziale na poszczególne tkanki i organy u ptaków koncentracja jonów wykazuje dość istotne zróżnicowanie. Przykładowo w tkance płynnej – krwi, stężenia pojedynczych pierwiastków w stanie równowagi fizjologicznej ustroju są charakterystyczne dla typu użytkowego drobiu, fazy wiekowej, co stanowi dość często w praktyce weterynaryjnej diagnostyki klinicznej ważny marker zaopatrzenia organizmu w te makroskładniki mineralne i równocześnie służyć może jako jeden z parametrów w szacowaniu potrzeb pokarmowych (Tab. 2) (Teeter i wsp., 1996).
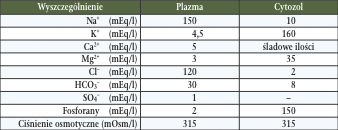
Jak podaje między innymi Larbier i wsp., (1995), Bączkowska (1987), Oviedo-Rondon i wsp., (2001) oraz Murakami i wsp., (2001) podział w zakresie funkcyjnym pomiędzy wymienionymi jonami jest stały w organizmie ptaków, a dynamiczne zmiany obserwowane są w rozkładzie ich stężeń (koncentracji) w komórkach i ich otoczeniu pod wpływem wielu czynników, przykładowo: temperatury otoczenia, szeroko rozumianej jakości żywienia (w tym zawartości sumy jonów w paszach) jak i jakości wody pitnej (zawartość w niej rozpuszczonych jonów). W poniższej tabeli przedstawiono rozkład jonów tj. anionów i kationów w plazmie i cytozolu podczas równowagi fizjologicznej w ustroju ptaków w mEq/l* (Tab. 3) (za Larbier i wsp., 1995).
Wspomniany już podział funkcyjny wymienionych makroelementów obejmuje ich status w utrzymywaniu odpowiedniego bilansu wody w organizmie i to na poziomie komórkowym jak i tkankowym (tkanki płynne), regulowanie ciśnienia osmotycznego, współuczestniczenie w utrzymaniu wyrównanego odczynu środowiska wewnętrznego i zewnętrznego poprzez układy buforujące (przeciw alkalozom i acidozom metabolicznym), w tworzeniu i utrzymywaniu bilansu elektrolitowego tzw. DEB (PBE) oraz współtworzenie struktur koloidów w komórkach, występowanie jako grupy czynne w aktywatorach i de aktywatorach wielu enzymów przemian komórkowych np. w cytozolu komórki.
Pozostałe funkcje elektrolitów w organizmie dotyczą też regulacji gazowej organizmu ptaków (np. jony potasu, chloru, sodu, węglanowe), utrzymania termogenezy (proces stałocieplności), regulacji i przebiegu transportu w tym dostarczania tlenu, regulowania i współdziałania z gospodarką hormonalną. Pełnią też one ważne funkcje strukturotwórcze, a więc wpływają na procesy rozwoju układu chrzęstno-kostnego. Również są regulatorami pracy układów np. nerwowego oraz pracy zespołów mięśni np. oddechowych (regulują częstotliwość oddechów i tym samym zdolność ochładzania organizmu), mięśnia sercowego, co wynika między innymi z przytoczonych już ww. funkcji.
Poniżej przedstawiono krótką indywidualną charakterystykę najistotniejszych jonów – elektrolitów u drobiu (Larbier i wsp., 1995; Hooge, 2003; Scott, 1978; Wertelecki 2003):
- Sód – jak wspomniano stanowi w warunkach równowagi fizjologicznej ptaków do 93% wszystkich jonów płynu otaczającego komórki. Znaczna część rezerw tego pierwiastka zgromadzona jest w tkance kostnej (około 1/3 całkowitej zawartości sodu w ciele ptaka) oraz wtórnie wynika z procesów reabsorpcji zwrotnej w nerkach. Sód zawarty w osoczu wywiera istotny wpływ na utrzymanie równowagi osmotycznej (w tym transportu cząsteczek wody) i jednocześnie ma wpływ na zachowanie prawidłowych, z punktu widzenia fizjologii organizmu, relacji – stosunków pomiędzy nim, a pozostałymi dwoma pierwiastkami: potasem i chlorem. Jon sodowy również może oddziaływać antagonistycznie na niektóre z enzymów komórkowych, które są aktywowane przez jony magnezu lub potasu, a dezaktywowane właśnie przez sód. Sód wraz z niżej wymienionymi elektrolitami pośrednio wpływa na rozwój tkanki kostnej oraz na stopień mineralizacji skorupy jaja.
- Potas jest jonem skoncentrowanym we wnętrzu komórek, a również nosi on miano czwartego makroelementu ustroju ptaków, a drugiego pod względem istotności w regulacjach bilansu kationów i anionów.
Koncentracja tego pierwiastka w krwinkach czerwonych ptaków może wynosić do 25 razy więcej niż w osoczu. Również specjalistyczne zespoły tkankowe zbudowane z komórek przewodzących, a więc układu nerwowego czy też komórki mięśni, w tym mięśnia sercowego, zawierać mogą do 20 razy więcej tego pierwiastka niż płyny międzykomórkowe je otaczające (Scott, 1978). Rozkład jonów potasu w organizmie ptaków jest dość stały w stanie równowagi fizjologicznej. W ustroju ptaków oprócz funkcji regulacji osmotycznej, bilansu elektrolitowego DEB (w tym regulacji równowagi kwasowo-zasadowej), aktywacji enzymów czy też wpływu na stopień metabolizmu związków azotowych (aminokwasów) i węglowodanów, potas w formie jonowej współuczestniczy też w przemianie innych makroelementów strukturalno-funkcyjnych, np. fosforu, co ma szczególne znaczenie w żywieniu kur nieśnych. Sód i potas wykazują silne działanie alkalogeniczne w organizmie ptaków.
- Chlor, w przeciwieństwie do ww. jonów jest jonem ujemnym, obecnym przede wszystkim w płynach zewnątrz komórkowych i wykazującym silne powinowactwo do sodu. Poprzez właściwości fizyko-chemiczne, określane mianem elektrolitowych stanowi ważny składnik w regulacji wzajemnych oddziaływań jonowych poza komórką, choć jest też transportowany do wnętrza komórek podobnie jak potas. Chlor spełnia istotną funkcję w regulacji stężenia jonów wodorowych w soku żołądkowym i uczestniczy w syntezie kwasu solnego. Ma to decydujący wpływ na procesy przemian tj. stopień trawienia białka pasz. Aczkolwiek w reakcjach biochemicznych zachodzących w organizmie sam wykazuje bardzo słabe powinowactwo do peptydów i polipeptydów. Z drugiej strony uznaje się chlor za składnik mineralny warunkujący biodostępność innych składników mineralnych tych, które mogą być absorbowane ze światła przewodu pokarmowego przy jego kwaśnym odczynie. Jon ten pośrednio wpływa na zmianę parametrów fizykochemicznych tkanki płynnej – krwi, np. zagęszczenie i na zmianę ciężaru właściwego, a także regulację bilansu wodnego organizmu w tym w procesie sekrecji wody na błony śluzowe w oddychaniu i chłodzeniu (parowanie). Współuczestniczy z omówionymi już jonami (Na i K) w procesie rozwoju tkanki chrzęstnej i to temu trójstronnemu oddziaływaniu przypisuje się znaczący wpływ na procesy jej kształtowania w interakcji z niektórymi aminokwasami (prolina, histydyna, lizyna, glicyna oraz seryna). Chlor jako jon przeciwny dla sodu i potasu wykazuje silne właściwości acidogenne.
Wzajemne oddziaływania omawianych jonów, tj. sodu, potasu i chloru stanowią o homeostazie organizmu oraz o tzw. gospodarce elektrolitowej, wodnej (bilans wodny) i mają wpływ na procesy termogenenzy i utrzymanie/regulacje stałocieplności.
Stadia oddziaływania wzajemnego jonów mogą przyjmować trzy podstawowe formy:
- równowagi (bilans zerowy, zrównoważony stan),
- nadmiaru (bilans dodatni dla danego jonu lub jonów, możliwość powstawania antogonizmów),
- niedoboru (bilans ujemny dla danego jonu lub jonów, możliwość powstawania antagonizmów lub synergizmu deficytowego).
W przypadku nadmiernego pobrania sodu u ptaków obserwuje się wzrost przemiany nerkowej prowadzący do próby zwiększenia wydalania tego pierwiastka z organizmu, a proces ten sprzężony jest z ponadnormatywnym spożyciem wody. Równocześnie ze wzrostem wydalania sodu obserwuje się ustanie reabsorpcji kanalikowej w nerkach. Niemniej w sytuacji, kiedy fizjologiczna zdolność wydalnicza nerek przestaje być wystarczającą dla ustroju dochodzi do zatrucia sodem: w badaniu sekcyjnym ptaków często obserwuje się opuchliznę nerkową (hypertrofia nerkowa), wole jest wypełnione płynem, wyraźne zmiany jakościowe we krwi. W obrazie subklinicznym w stadzie widoczne są ptaki z wysiękiem śluzowatym z otworów nosowych, obserwuje się też przyhamowanie wzrostu, spadek spożycia paszy, wyraźną nerwowość – pobudliwość, skupianie się ptaków, rozpostarcie skrzydeł, odwodnienie organizmu (biegunki) i wzrastające upadki.
Przeciwstawny obraz można zaobserwować u drobiu przy utrzymujących się dłuższy czas stanach niedoborowych sodu: zahamowanie wzrostu i rozwoju, spadek aktywności ruchowej w tym przy pobieraniu paszy, obniżenie stopnia przemian metabolicznych aminokwasów, energii i węglowodanów, zmiany strukturalne w kościach długich, keratynizacja rogówki, zmiany w gonadach, zmniejszenie objętości osocza, wzrost poziomu kwasu moczowego we krwi, zaburzenia równowagi, aż do wystąpienia wstrząsów – neuro-porażeń, a nawet kanibalizm (Larbier i wsp. 1995, Scott, 1978, Wertelecki, 2003). Organizm ptaków przystosował się w toku ewolucji dość dobrze do sytuacji niskiej podaży tego makroskładnika w pokarmie.
Nadmiar jonów potasu jest nieco mniej szkodliwy dla organizmu ptaków niż jonów sodu (do pewnej granicy tolerancyjności). Koncentracja potasu jest płynnie regulowana przez wydalanie nerkowe (w głównej mierze) wraz z interakcją z sodem, o czym już wspomniano. Niemniej w sytuacjach wyjątkowych, gdy podaż tego składnika przekracza znacząco normatywne poziomy dawkowania (rekomendacje), wówczas podobnie jak przy sodzie nerki nie są w stanie sprostać jego wydalaniu, szczególnie potasu plazmatycznego. Dochodzi wówczas do zaburzeń pracy serca, zmiany wodnistości odchodów, zaburzeń osmotycznych i kwasowo-zasadowych, obniżenia niektórych przemian metabolicznych, a w konsekwencji doprowadzić to może do śmierci ptaków. Nadmiar potasu powoduje wzrost reabsorpcji sodu (Larbier i wsp., 1995).
Niedobory potasu, to bardzo rzadkie przypadki, noszą nazwę hipokalcemii – wewnątrz komórek następuje obniżenie stężenia jonów potasu, co prowadzić może do zakwaszenia środowiska wewnętrznego komórki. Głównymi objawami niedoboru jonów K są: osłabienie mięśni, zmniejszenie napięcia ścian jelit, aż do ich rozciągnięcia, osłabienie pracy serca i mięśni oddechowych. Obniżenie kilkuprocentowe poziomu potasu plazmatycznego w okresie wysokich temperatur u ptaków wyraźnie potęguje wzrost ich śmiertelności w tym okresie, a szczególnie to jest widoczne u młodych przegrzanych piskląt lub ptaków o wysokiej masie ciała (przed okresem uboju).
Zaburzenia gospodarki jonów chloru z reguły wykazują formę nadmiaru, niemniej w przypadkach niedoborowych, szczególnie przy skarmianiu pasz zawierających, poniżej 0,7 g Cl/kg, obserwuje się u ptaków obniżenie tempa wzrostu, zagęszczenie fizyczne krwi i płynów śródtkankowych, odwodnienie ustroju wraz z radykalnym spadkiem chlorków we krwi. Mogą występować przy dłuższym deficycie objawy neurologiczne – porażenia ptaków, pozycja z wyciągniętymi kończynami.
Złożoność interakcji (współoddziaływania) pomiędzy omawianymi jonami w organizmie winna być zawsze przedmiotem szerokiej i zbiorczej analizy przy optymalizacji mieszanek dla drobiu. Przykładem takiej interakcji jest omówione już oddziaływanie sodu i potasu w zakresie wyrównywania ich stężeń w płynach ustrojowych, a także układ regulujący transport komórkowy noszący potoczną nazwę pompy sodowo-potasowej, złożonej z jonów sodu, potasu i współuczestnictwa ATP i ATPazy. Pompa ta usuwa na zewnątrz komórki sód, a przenosi do jej wnętrza jony potasu (w stanie równowagi fizjologicznej). Podstawowym źródłem energii w tym procesie jest ATP powstające z komórkowego utleniania glukozy (Larbier i wsp., 1995). Zaś najważniejszym medium jest – faza wodna i cząsteczki wody.
Tak więc reasumując powyższe podsumowanie fizjologicznej roli jonów, ich związku z procesami metabolicznymi i bilansem wody – utożsamianym ze zdolnością do utrzymania i regulacji stałej temperatury ciała, stwierdzić należy, że całość tych procesów winna być brana pod uwagę, gdy chce się regulować, wspomagać lub ukierunkowywać homeostazę termiczną i bilans wodny organizmu.
dr inż. Tomasz Wertelecki, mgr inż. Piotr Łambucki