


Diana Siebert 1*, Daulat Rehman Khan 1, David Torrallardona 2
1 CJ Europe GmbH, 60549 Frankfurt am Main, Germany; Ten adres pocztowy jest chroniony przed spamowaniem. Aby go zobaczyć, konieczne jest włączenie w przeglądarce obsługi JavaScript.
2 Institute of Agrifood Research & Technology (IRTA), 43120 Constantí (Tarragona), Spain; Ten adres pocztowy jest chroniony przed spamowaniem. Aby go zobaczyć, konieczne jest włączenie w przeglądarce obsługi JavaScript.
* Kontakt: Ten adres pocztowy jest chroniony przed spamowaniem. Aby go zobaczyć, konieczne jest włączenie w przeglądarce obsługi JavaScript., tel. +49-6929-9144-271
https://doi.org/10.3390/ani11051255
Jaki powinien być stosunek waliny do lizyny, w paszy do uzyskania optymalnych parametrów użytkowych u odsadzonych prosiąt?
Istnieje kilka przyczyn stojących za tendencją do obniżania zawartości białka surowego (CP) w paszach dla świń. Z jednej strony rośnie globalny trend obniżania nadmiaru azotu w paszach, który jest wydalany z moczem i kałem oraz jego negatywny wpływ na środowisko [1]. Z drugiej strony, względy ekonomiczne mogą być również motorem napędowym w kierunku pasz niskobiałkowych, zwłaszcza gdy ceny śruty sojowej są wysokie, a białko staje się jednym z najdroższych składników pokarmowych. Ponadto, nadmiar białka, zwłaszcza u prosiąt odsadzonych wiąże się z zaburzeniami żołądkowo-jelitowymi, takimi jak biegunka poodsadzeniowa [2-4]. W jednym z ostatnich opracowań podkreślono dodatkowo, że zmiany w poziomie białka w paszy wpływają na regulację wielu genów [5], chociaż istnieje potrzeba większej ilości badań, aby ocenić związek pomiędzy zróżnicowaną zawartością aminokwasów a epigenetycznym zmianami u świń. W przypadku pasz o niskiej zawartości CP, udział krystalicznych aminokwasów uzupełniających (AA) wzrasta proporcjonalnie, aby zaspokoić zapotrzebowanie zwierząt na aminokwasy egzogenne i utrzymać optymalne wyniki wydajności.
Komercyjne pasze dla świń są rutynowo uzupełniane lizyną (Lys), metioniną (Met), treoniną (Thr) i tryptofanem (Trp). W paszach dla trzody opartych na roślinnych źródłach białka walina (Wal) powinna być piątym ograniczającym aminokwasem (przed izoleucyną) [6-8]. W przeszłości, stosowanie krystalicznej Wal nie było powszechne z powodu jej ograniczonej dostępności i wysokiej ceny, jednak w ostatnich latach sytuacja ta uległa zmianie. Bilansowanie zapotrzebowania zwierząt na AA i uniknięcie nadmiernej podaży lub niepożądanych interakcji wymaga dokładnej wiedzy na temat zapotrzebowania zwierzęcia na AA.
Przeprowadzono różne badania w celu zbadania optymalnego stosunku Met, Thr i Trp do Lys u świń. W przeciwieństwie do tego, optymalny stosunek Wal do Lys został zbadany w niewielkim stopniu, mimo, że dostępna literatura wskazuje wyraźnie, że niedobór Wal prowadzi do zmniejszenia pobierania paszy i w konsekwencji do słabych wyników produkcyjnych u warchlaków [9,10]. Kilka czynników, takich jak płeć, etap produkcji czy stan zdrowia, może wpływać na to zapotrzebowanie.
Ponieważ Wal, leucyna (Leu) i izoleucyna (Ile) należą do grupy aminokwasów rozgałęzionych (BCAA) mają wspólny katabolizm [11], interakcje żywieniowe pomiędzy tymi aminokwasami mogą potencjalnie wpływać na zapotrzebowanie na ten aminokwas [12]. Rzeczywiście, istnieją sprzeczne informacje na temat zapotrzebowania na Wal u prosiąt od 5 do 25 kg w dostępnej literaturze. National Research Council (NRC 2012) zaleca, aby dWal : dLys wynosił 0,63 [13], podczas gdy inni podają, że dWal : dLys wynosi. 0,63-0,65 [14] lub 0,70 [15,16].
Dodatkowo, różne modele statystyczne stosowane do oszacowania zapotrzebowania mogą skutkować zróżnicowanymi zaleceniami. Celem niniejszej pracy było określenie zapotrzebowania dWal: dLys u prosiąt po odsadzeniu przy zastosowaniu różnych modeli regresji.
MATERIAŁY I METODY
Praca została przeprowadzona w gospodarstwie doświadczalnym instytutu Agrifood Research & Technology (IRTA) po uzyskaniu zgody komitetu etycznego IRTA. Research & Technology (IRTA) po zatwierdzeniu przez Komitet Etyczny ds. Experimentation (CEEA); kod badania P-531, kod uchwały E-21/2021.
1. Zwierzęta i pomieszczenia
Przeprowadzono 43-dniową próbę żywieniową z udziałem ogółem dwustu odsadzonych prosiąt
((Duroc x Landrace) x Pietrain; różnej płci). Prosięta zostały pozyskane ze stada loch (IRTA, Hiszpania) w wieku około 31 dnia życia (5 dni po odsadzeniu).
Na początku testu (dzień 0) średnia początkowa masa ciała prosiąt wynosiła 8,7 kg ± (1.1). Prosięta zostały losowo podzielone według początkowej masy ciała na dziesięć bloków, a każdy blok składał się z pięciu kojców z czterema prosiętami w każdym. Prosięta były w ten sposób utrzymywane w 50 kojcach w trzech pomieszczeniach do odsadzania (odpowiednio 20, 20 i 10 kojców na pomieszczenie). Pomieszczenia były wyposażone w automatyczne ogrzewanie, wymuszoną wentylację i podłogi rusztowe, a temperatura była regulowana tak, aby stopniowo obniżała się z 30°C do 24°C w ciągu pierwszych 14 dni po odsadzeniu oraz z 24°C do 21°C od 14 do 48 dnia po odsadzeniu. Pomieszczenia wyposażono w świetlówki rurowe i zaprogramowano cykl 16 : 8 godz. światło: ciemność. Wszystkie prosięta były codziennie monitorowane pod kątem nieprawidłowości takich jak: klinicznych objawów chorób, nieprawidłowego zachowania i śmiertelności przez cały okres trwania doświadczenia.
2. Projekt doświadczenia i program żywieniowy
Wszystkie prosięta odsadzono w 26 dniu życia i przez pięć kolejnych dni otrzymywały tę samą komercyjną paszę po odsadzeniu. Następnie (dzień 0 testu), prosięta zostały podzielone według masy ciała i przydzielone do grup testowych w losowo wybranym układzie bloków z 10 blokami i pięcioma mieszankami testowymi. Kojce w każdym bloku były umieszczone obok siebie i w tym samym pomieszczeniu. Masa ciała poszczególnych świń i pobranie paszy przez kojec były kontrolowane na początku doświadczenia (dzień 0), w dniu 13, w dniu 29 i na końcu doświadczenia (dzień 43). Średni dzienny przyrost masy ciała (ADG), średnie dzienne pobranie paszy (ADFI), i współczynnik wykorzystania paszy (FCR) obliczono dla każdego okresu doświadczalnego, jak również dla całego okresu trwania testu.
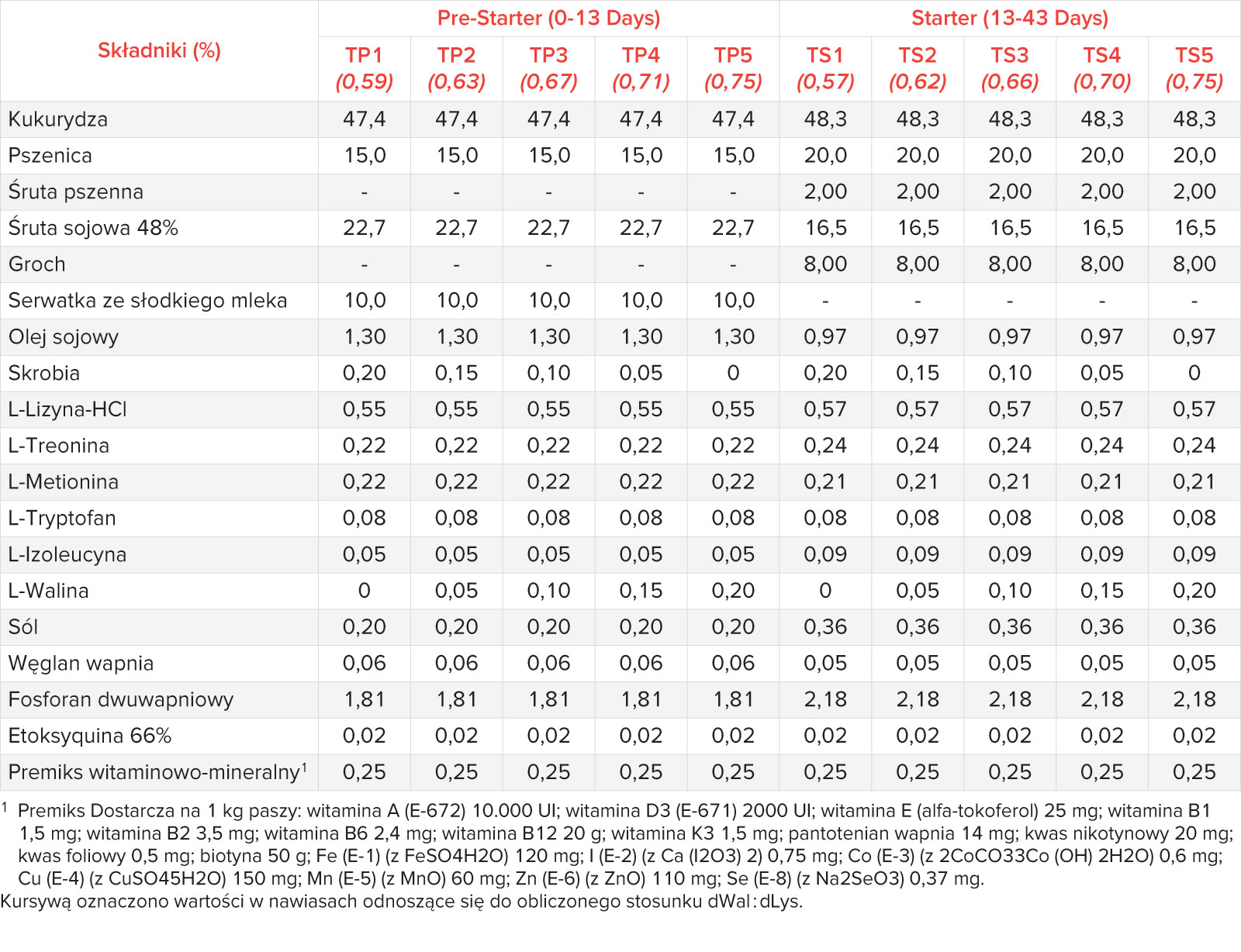
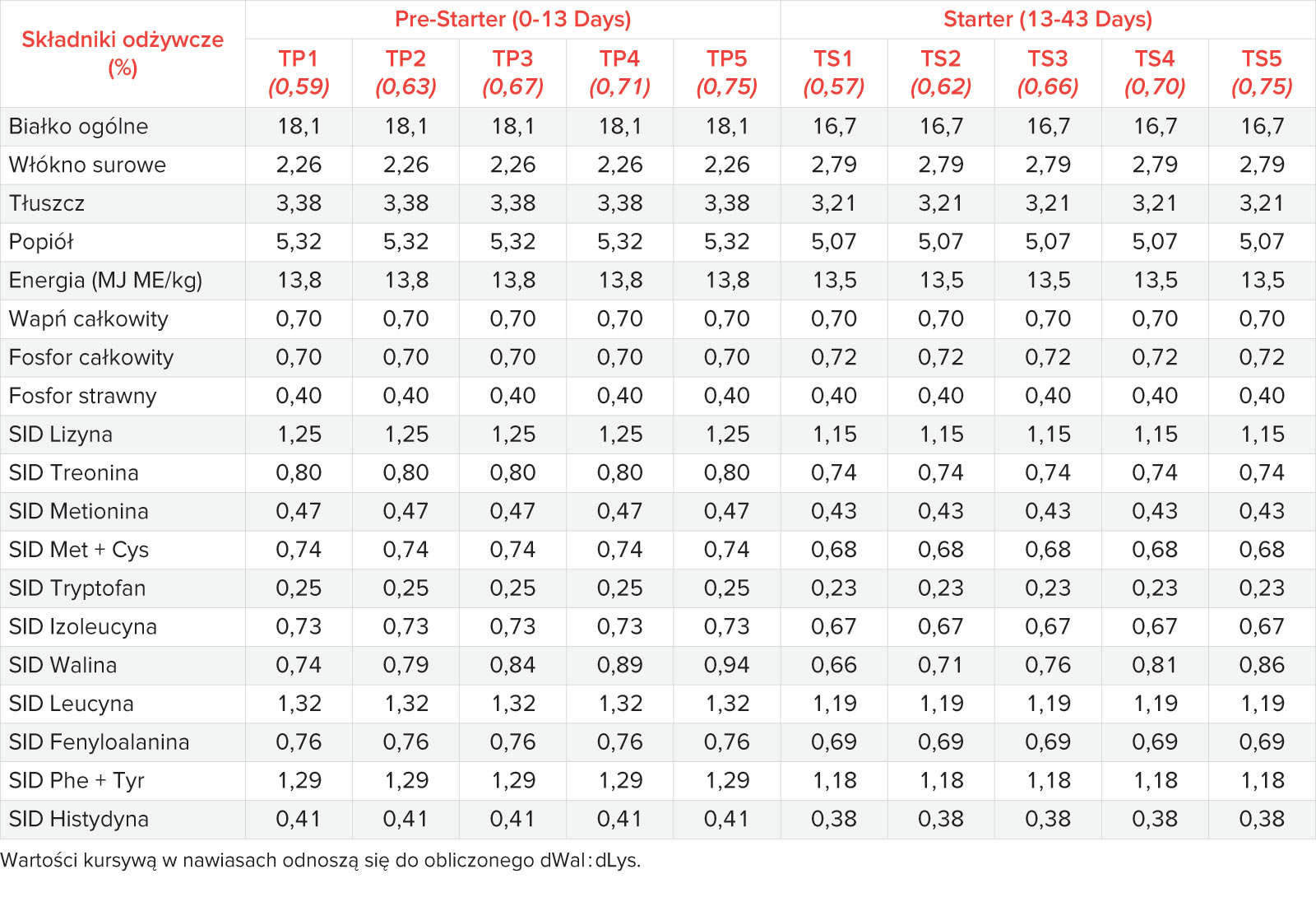
Mieszanki składały się głównie z kukurydzy, pszenicy i śruty sojowej, a także z aminokwasów, witamin i składników mineralnych (Tab. 1). Zawartość w diecie wszystkich niezbędnych aminokwasów, z wyjątkiem waliny, utrzymywano na stałym poziomie. Wzrastające ilości Wal (BESTAMINO™ L-Valine, CJ Europe GmbH, Frankfurt am Main, Niemcy) w poszczególnych grupach żywieniowych osiągnięto poprzez zastąpienie skrobi kukurydzianej krystaliczną waliną. Pasza została wyprodukowana w doświadczalnej wytwórni pasz IRTA w formie granulatu (3 mm) i podawana ad libitum. Program żywienia podzielono na dwie fazy żywienia: pre-starter (13,8 MJ ME, 12,5 g/kg SID Lizyny) od 0 do 13 dnia testu i diety starter (13,5 MJ ME, 11,5 g/kg SID Lizyny) pomiędzy 13-43 dniem testu (Tab. 2). Stosunek dWal : dLys ustalono na poziomie 0,59; 0,63; 0,67; 0,71 i 0,75 w pięciu paszach typu pre-starter (TP1 do TP5) oraz 0,57; 0,62; 0,66; 0,70 i 0,75 w pięciu paszach typu starter (TS1 do TS5). Wartości strawności są standaryzowanym współczynnikiem strawności jelitowej (SID) zaczerpnięte z opublikowanych tabel [17].
3. Analiza pasz
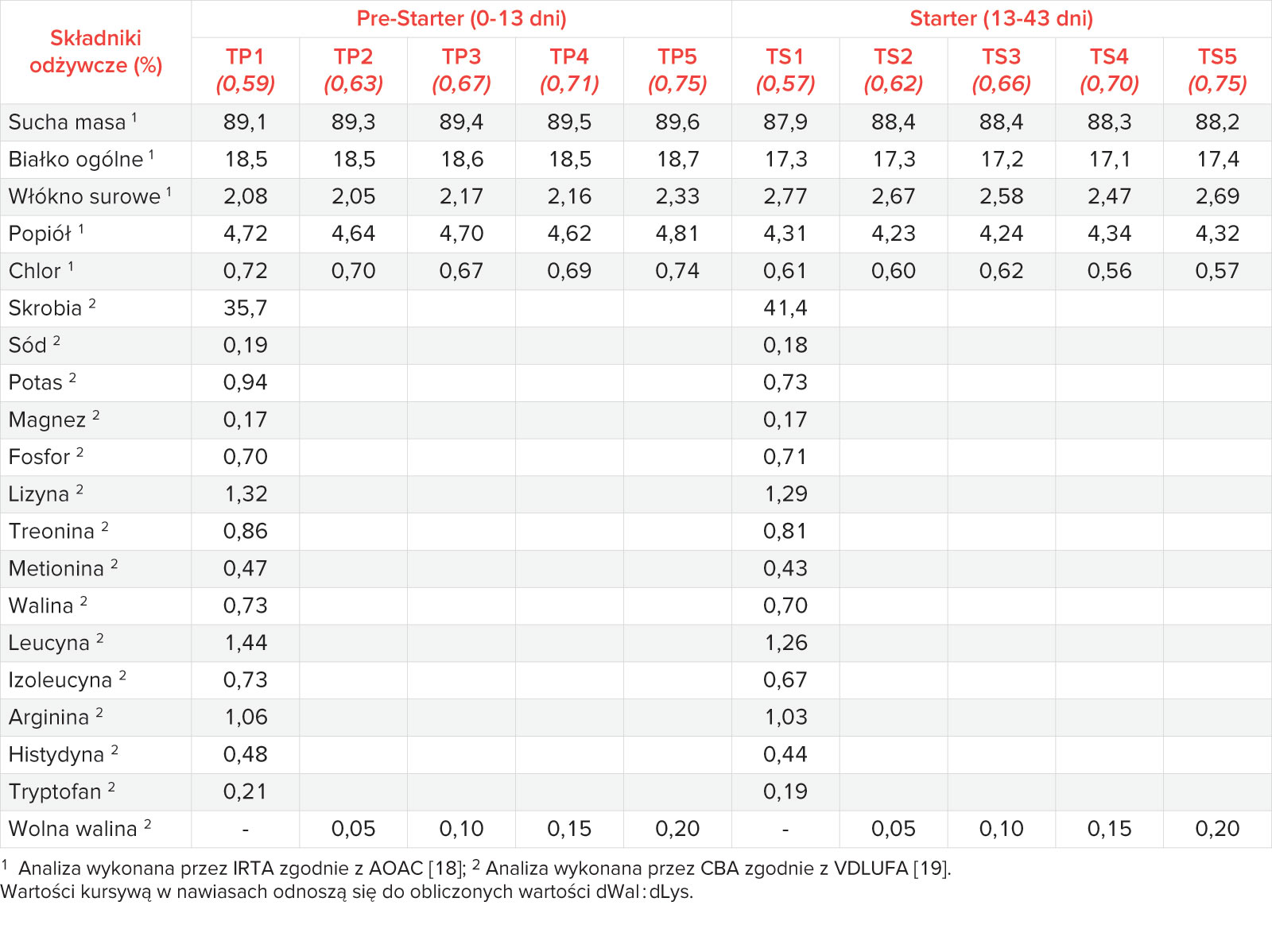
Wszystkie pasze analizowano pod kątem ogólnej zawartości składników odżywczych zgodnie z oficjalnymi metodami AOAC [18]. Analizę wolnej (dodanej) waliny przeprowadzono zgodnie z oficjalnymi metodami VDLUFA [19]. Obliczone i analizowane wartości odżywcze pokrywały się ze sobą w dobrym stopniu (Tabela 3).
4. Analiza statystyczna
Dane sprawdzono pod kątem normalności i homoscedastyczności, a zmierzone parametry porównywano pomiędzy poszczególnymi grupami metodą ANOVA z wykorzystaniem procedury GLM pakietu statystycznego SAS w wersji 9.4 (SAS Institute Inc., Cary, NC, USA), a jako jednostkę doświadczalną przyjęto kojec. Efekty bloku (grupa o początkowej masie ciała i lokalizacja kojca) oraz paszy uwzględniono w modelu. Średnie najmniejszych kwadratów, prawdopodobieństwa różnic oraz błędy standardowe średniej zostały obliczone w celu oceny różnic między średnimi pasz. Dane w tabelach są przedstawione jako średnie najmniejszych kwadratów. Kontrast ortogonalny został również użyty do określenia liniowych i kwadratowych odpowiedzi na rosnące stężenie Wal. Wszystkie stwierdzenia istotności statystycznej były oparte na poziomie p < 0,05.
W celu określenia optymalnego stosunku dWal : dLys, dane analizowano za pomocą dwóch nieliniowych modeli regresji przy użyciu procedury NLIN w SAS. Dwa zastosowane modele były następujące:
Model 1:
wykładniczy asymptotyczny (EA)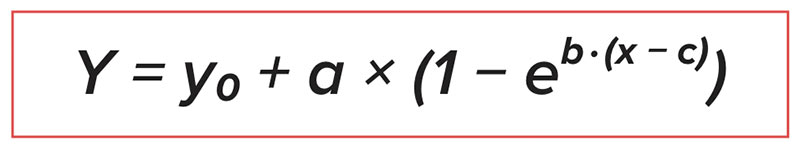 gdzie:
gdzie:
Y = zmienna odpowiedzi (masa ciała, średni dzienny przyrost, średnie dzienne pobranie paszy, współczynnik konwersji paszy);
x = stosunek dWal do dLys;
y0 = odpowiedź przechwytująca przy 0 poziomie uzupełniającej L-waliny (tj. stosunek dWal: dLys z paszy kontrolnej);
c = stosunek dWal: dLys dla paszy kontrolnej;
a = maksymalna odpowiedź na suplementację L-waliną (w stosunku do paszy kontrolnej); oraz
b = parametr modelu, który ma być oszacowany.
Maksymalna odpowiedź była analizowana jako 95% i 99% plateau.
Model 2:
Regresji krzywoliniowej-plateau (CLP) gdzie:
gdzie:
Y < R; Y = a dla x ≥ R; Y = zmienna odpowiedzi (masa ciała, średni dzienny przyrost, średnie dzienne spożycie paszy, współczynnik konwersji paszy);
x = stosunek dWal do dLys;
a = maksymalna odpowiedź na suplementację L-waliną (w stosunku do paszy kontrolnej);
R = zapotrzebowanie; oraz
U = parametry modelu, który ma być oszacowany.
WYNIKI
Próba przebiegła bez komplikacji, a więc nie było konieczności stosowania antybiotykoterapii podczas doświadczenia. Odnotowano niską śmiertelność. Trzy prosięta padły podczas fazy pre-starter (jedno należące do grupy TP1, jedno należące do grupy TP2 i jedno należące do grupy TP4). Dwa prosięta padły w fazie starter (należące do grupy TS4) z klinicznymi objawami choroby układu oddechowego. Dodatkowo, dwa prosięta zostały ubite w 29 dniu z powodu bardzo słabej wydajności (należące do grupy TS1). Dane tych zwierząt, wraz z danymi dodatkowych sześciu zwierząt, które zostały zidentyfikowane jako wartości odstające za pomocą testu Smirnoffa-Grubbsa [20], nie zostały wykorzystane do obliczeń. Ich spożycie paszy zostało oszacowane na podstawie spożycia paszy w ich kojcach do czasu ich usunięcia, przyrostu ich masy ciała oraz przyrostu masy ciała ich kolegów z kojca, według Lindemann i Kim (2006) [21].
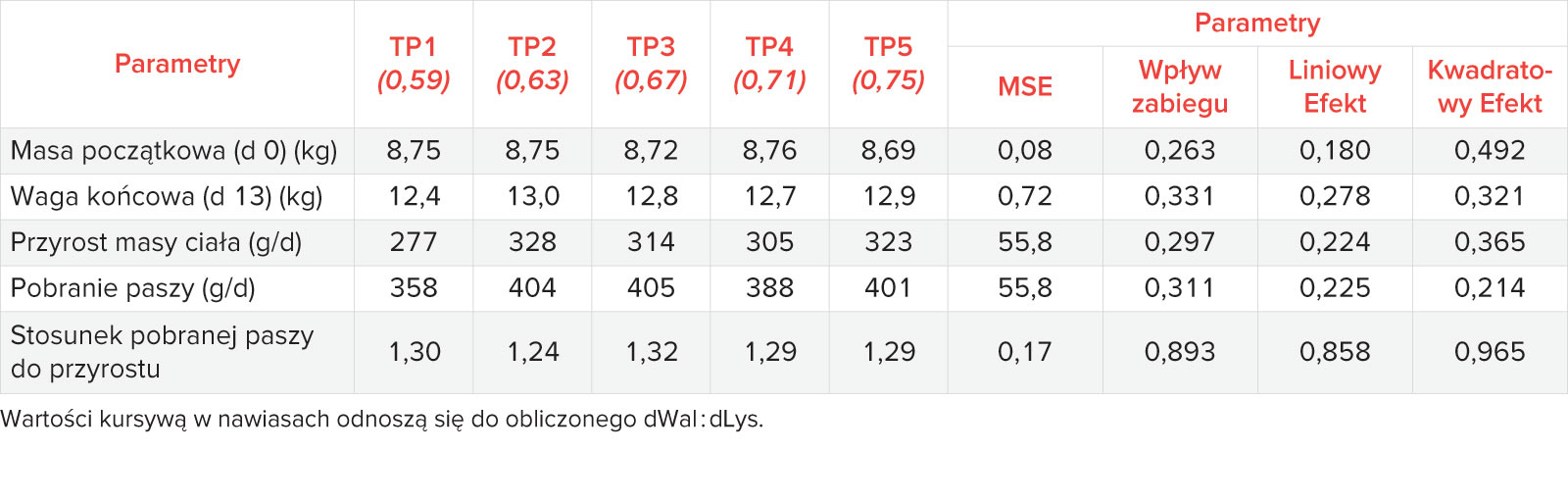
1. Przyrost masy ciała
Dane dotyczące przyrostu masy ciała w fazie pre-starter nie pasowały do żadnego z modeli regresji. Przyrosty masy ciała mieściły się w przedziale od 277 g/d do 328 g/d i nie stwierdzono statystycznie istotnych różnic pomiędzy poszczególnymi grupami (Tabela 4). W fazie starter oszacowane optymalne dWal : dLys wynosiły 0,68 (95% maksymalnej odpowiedzi) i 0,74 (99% maksymalnej odpowiedzi) w oparciu o użycie modelu EA. Wykorzystując model CLP dla tego samego okresu, szacowany stosunek dWal : dLys wynosił 0,67. Oceniając całkowity okres testu (d 0-43), szacowane współczynniki dWal : dLys dla przyrostu masy ciała wynosiły 0,65 (95% maksymalnej odpowiedzi) i 0,70 (99% maksymalnej odpowiedzi) przy użyciu modelu EA i 0,65 przy modelu CLP (Tabela 5).
2. Pobranie paszy
Liczbowo, najniższe pobranie paszy w fazie pre-starter obserwowano w przypadku paszy TP1; nie stwierdzono jednak różnic statystycznych pomiędzy poszczególnymi grupami (Tab. 4). W fazie startowej prosięta żywione paszą TS1 zjadły istotnie mniej paszy niż prosięta w pozostałych grupach suplementowanych Wal (p < 0,05; tab. 6), a nie stwierdzono statystycznie istotnych (p > 0,05) różnic wśród grup suplementowanych Wal (TS2-TS5).
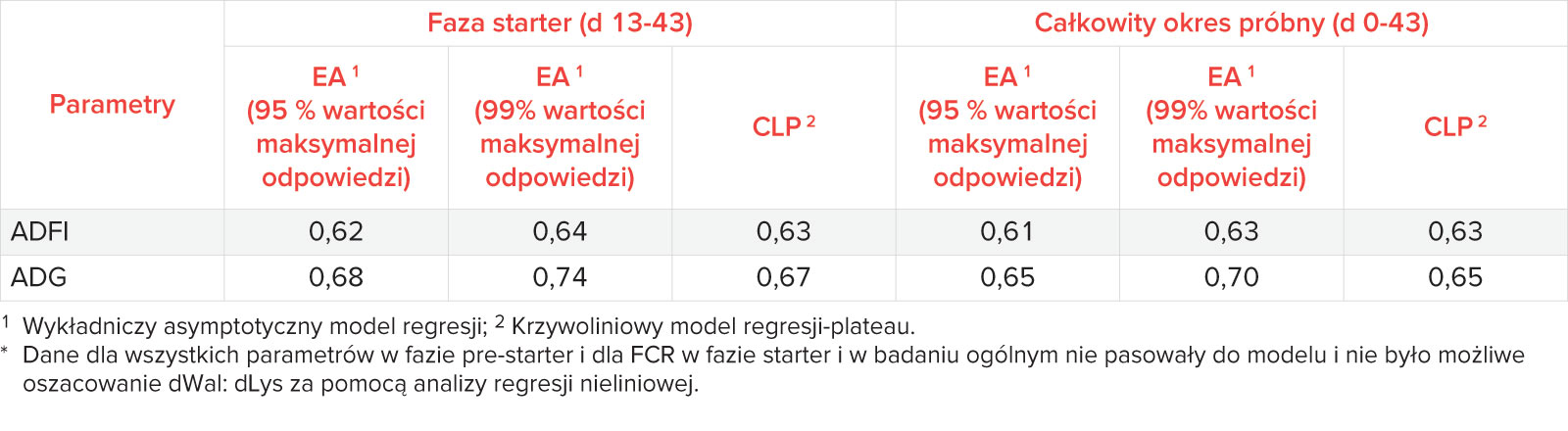
W przypadku przyrostu masy ciała, w fazie pre-starter, nie było możliwe oszacowanie optymalnego dWal : dLys za pomocą modeli regresji. Jednakże, w fazie starter, oszacowane dWal : dLys wynosił 0,62 (95% maksymalnej odpowiedzi) i 0,64 (99% maksymalnej odpowiedzi) w oparciu o użycie modelu EA, a także 0,64 (99% maksymalnej odpowiedzi) w oparciu o użycie modelu EA oraz 0,63 przy zastosowaniu modelu CLP. Biorąc pod uwagę całkowity okres testu (d 0-43), optymalne dWal : dLys dla spożycia paszy zostały ustalone na poziomie 0,61 (95% maksymalnej odpowiedzi) i 0,63 (99% maksymalnej odpowiedzi) w oparciu o użycie modelu EA oraz 0,63 w oparciu o model CLP (Tabela 5).
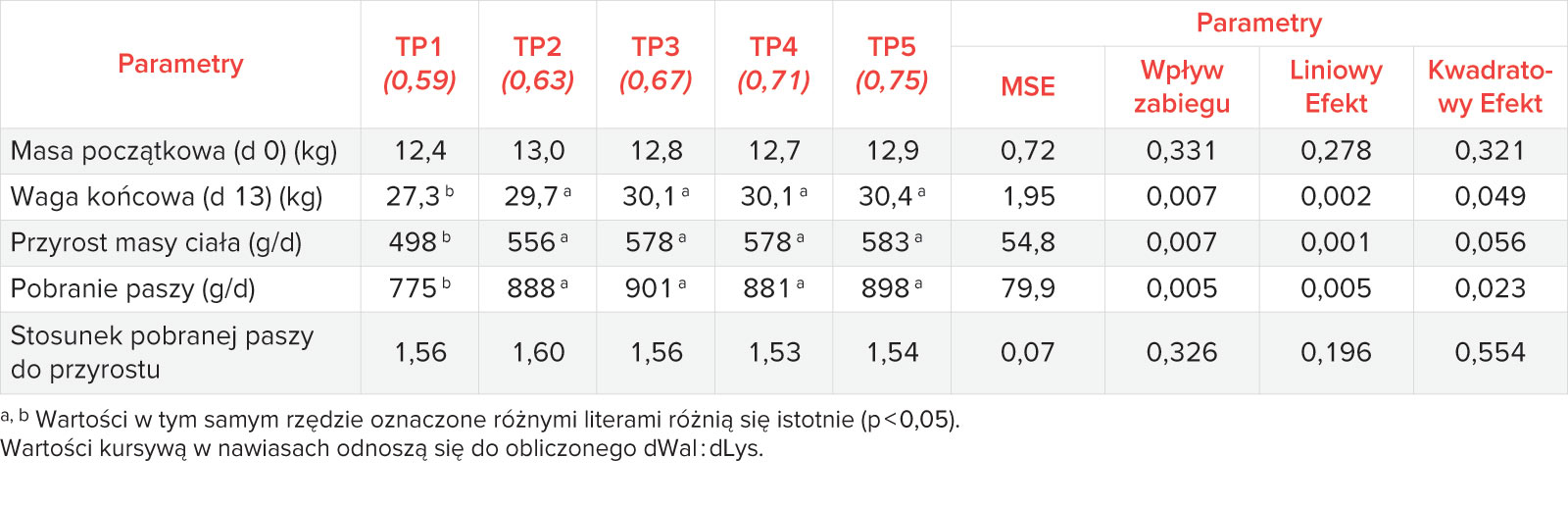
3. FCR
FCR wahał się między 1,24 a 1,30 dla fazy pre-starter (Tabela 4), między 1,53-1,66 dla fazy starter (Tabela 6) i między 1,48-1,52 dla całego testu (Tabela 7). Odpowiedzi na dodatek L-waliny na efektywność wykorzystania paszy nie były istotne dla żadnej z faz testu.
4. Dyskusja
Celem badań było określenie zapotrzebowania na dWal : dLys u prosiąt po odsadzeniu dla uzyskania optymalnej wydajności. W tym badaniu, przyrost był znacznie zmniejszony po zastosowaniu paszy podstawowej z niedoborem Wal, co dowodzi, że niedobór Wal ma wpływ na opóźnienie wzrostu [22]. Niedobory lub brak równowagi pomiędzy aminokwasami rozgałęzionymi (BCAA) w paszy zostały przedstawione jako powód zmniejszonego pobrania paszy przez kilku autorów. W teście podwójnego wyboru Suarez i in, (2012) [23] przetestowali trzy różne poziomy dodatku Wal (0,73; 0,78 i 0,83 Wal: Lys) oraz dietę bez krystalicznej Wal [23]. Najmniejszy stopień preferencji pobrania paszy występował w mieszankach bez dodatku Wal, natomiast pasze z dodatkiem Wal były preferowane niezależnie od poziomu Wal. Dane uzyskane w modelu mysim sugerują, że podwzgórzowa somatostatyna może być związana z dietą z niedoborem Wal jako centralnego czynnika prowadzącego do anoreksji [24]. Podanie szczurom diety z niedoborem Wal prowadziło do ciężkiej anoreksji, w tym spadku stężenia Wal w płynie mózgowo-rdzeniowym, jak również hiper-ghrelinemii [25]. Centralne podanie greliny zwiększało spożycie diety z niedoborem Wal, natomiast w przypadku aplikacji systemowej nie zaobserwowano żadnego efektu [25]. W niniejszych badaniach pasza podstawowa z niedoborem Wal miała najniższe pobranie. Wśród poszczególnych grup suplementowanych Wal nie zaobserwowano różnic statystycznych w pobraniu paszy.
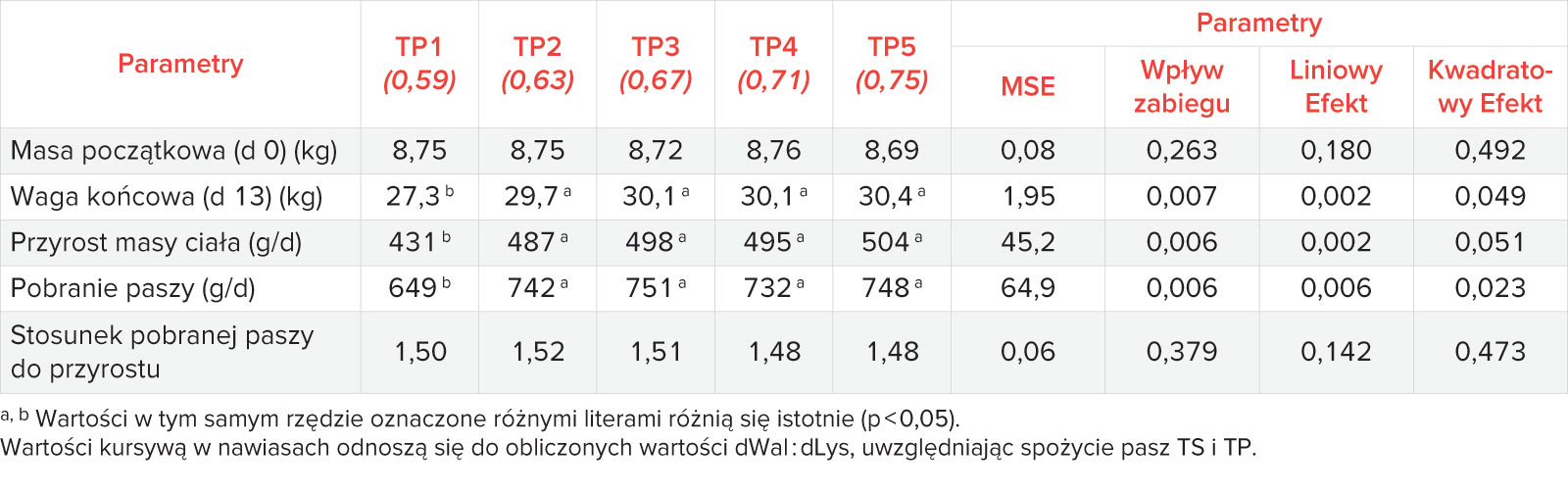
Nie stwierdzono dodatkowej poprawy pobrania paszy przy zwiększeniu udziału Wal. Używając pobranie paszy jako kryterium odpowiedzi, szacowane zapotrzebowanie wynosiło odpowiednio 0,61; 0,63; 0,63 (model EA: 95% maksymalnej odpowiedzi, model EA: 99% maksymalnej odpowiedzi i model CLP) dla całkowitego okresu trwania testu. NRC zaleca optymalny stosunek 0,63 [13], jednak w większości badań nad dawką waliny obserwowano wyższe wartości optymalizujące ADFI. W badaniu typu dawka-odpowiedź u prosiąt o masie ciała od 8 do 14 kg [9], najniższe pobranie paszy stwierdzono również w przypadku 0,58 dWal : dLys, która była paszą z niedoborem Waliny. Autorzy ci również donoszą o wzroście pobrania paszy wraz z większym dWal : dLys, a optymalne dWal : dLys dla ADFI określono na poziomie 0,70, czyli wyższym niż w obecnych badaniach. Barea i in., (2009) podali nawet wyższy poziom wymagań dla optymalizacji ADFI (tj. 0,74 przy modelu liniowo-plateu i 0,81 przy modelu krzywoliniowo-plateu) [10]. Nie było jasne, dlaczego zapotrzebowanie na Wal dla ADFI było porównywalnie niskie i nie reagowało w sposób zależny od dawki, jak tego oczekiwano. Niedobory w zawartości BCAA w diecie zostały opisane przez wielu autorów jako mające negatywny wpływ na pobranie paszy. Obniżenie ADFI zaobserwowano w dietach z niedoborem Wal [8,10], ale także w dietach z niedoborem Ile [9]. Nadmiar Leu w połączeniu z niskim poziomem Wal w diecie miał negatywny wpływ na pobranie paszy i w konsekwencji na wydajność prosiąt [26,27]. BCAA są degradowane przez te same kompleksy układów enzymatycznych, dlatego nadmiar Leu może wpływać na zapotrzebowanie na Wal i Ile [12], ponieważ nadmiar Leu aktywuje degradację Wal i Ile. W tym badaniu, pasze zostały sformułowane tak, aby spełnić SID Leu : Lys na poziomie około 1,06 w pre-starterze i około 1,03 w fazie starter, natomiast SID Ile: Lys wynosił 0,58 w obu fazach. Ponieważ poziomy Leu są zbliżone do zaleceń NRC [13] (1,00), wydaje się mało prawdopodobne, że niski wskaźnik ADFI był spowodowany antagonizmem pomiędzy BCAA.
Chociaż nie zaobserwowano znaczących różnic w wydajności podczas fazy pre-starter, to jednak 0,59 dWal: dLys skutkowało najniższym przyrostem masy ciała i pobraniem paszy z liczbowego punktu widzenia, co sugeruje, że ta mieszanka mogła być nadal niedoborowa. Byłoby to zgodne z opinią Soumeh i in., (2015) [9], którzy zaobserwowali istotnie obniżoną wydajność u prosiąt w przedziale 8-14 kg przy zawartości 0,58 dWal : dLys. Brak statystycznej istotności w naszym badaniu może być spowodowany większą zmiennością danych w ciągu pierwszych kilku tygodni po odsadzeniu. Niemniej jednak, gdy wzięto pod uwagę cały okres testowy, stało się jasne, że diety 0,59 i 0,57 dWal : dLys były niedoborowe pod względem zawartości Waliny. Szacowane zapotrzebowanie 0,68 (model EA: 95% maksymalnej odpowiedzi) dla optymalnego wzrostu odpowiadało ogólnie wartości zaproponowanej przez Changa i Baker (1992) [28]. Nørgaard i Fernández (2009) testowali dodatek Wal (0,72 dWal : dLys) i Ile (0,61 dIle : dLys) lub ich kombinacji z dietą dla prosiąt, która była uboga w oba aminokwasy (0,61 dWal : dLys; 0,53 dIle : dLys) [29]. Dodatek Wal lub Wal i Ile do deficytowej paszy podstawowej istotnie poprawiło ADG odpowiednio o 14,8 i 19,4%. Tym samym Wal ograniczała wydajność zwierząt przed Ile [29], dowodząc, że Wal jest ważna w formułowaniu pasz dla prosiąt. W tym badaniu, ADG fazy pre-starter (prosięta 8-12 kg) wahało się między 277 a 328 g/d, podczas gdy Soumeh i in., (2015) [9] obserwowali zakres między. 304 g/d a 428 g/d u prosiąt o masie ciała 8-14 kg. Różnica w liczbach bezwzględnych może być wyjaśniona przez porównywalnie niższe spożycie paszy w tym teście.
Zapotrzebowanie na dWal : dLys dla maksymalnego ADG wynosiło 0,68; 0,74 i 0,67 w fazie starter (model EA: 95% maksymalnej odpowiedzi, model EA: 99% maksymalnej odpowiedzi i modelu CLP). Soumeh i in., (2015) [9] oszacowali zapotrzebowanie dWal : dLys na poziomie 0,67 (model linii łamanej) i 0,71 (model krzywoliniowy plateau), aby zmaksymalizować ADG. Wykorzystując model linear-plateau Barea i in. (2009) podali 0,70 dla maksymalnych przyrostów i 0,75 przy zastosowaniu modelu krzywoliniowego plateau. Gloaguen i in., (2011) wykazali średnie zapotrzebowanie dWal : dLys wynoszące 0,72; jednakże, obserwacja ta została dokonana w przypadku paszy o wysokiej zawartości Leu (165% Leu : Lys) [15], co potencjalnie wpływa na zapotrzebowanie z powodu wzajemnego oddziaływania BCAA. Z drugiej strony, literatura podaje również niższe wymagania dWal : dLys niż te zaobserwowane w niniejszej pracy. Gains i in., (2011) zasugerowali, że 0,65 dWal : dLys jest odpowiednie dla prosiąt o wadze 13-32 kg [30]. W niedawnym badaniu z udziałem 280 świń w wieku okołoodsadzeniowym zapotrzebowanie zostało oszacowane na około 0,63 w celu optymalizacji ADG, przy zastosowaniu modelu liniowego z linią łamaną [31]. W doświadczeniu z udziałem tuczników o masie ciała od 26 do 46 kg, Liu i in., (2015) wykazali. istotny wpływ modelu statystycznego na szacowane zapotrzebowanie [32]. Stosując model liniowy łamany, szacowane dWal : dLys dla maksymalizacji ADG wynosiło 0,62, podczas gdy wynosiło 0,71 przy zastosowaniu modelu kwadratowego [32]. Jako ogólna tendencja, modele EA mogą zawyżać zapotrzebowanie na dWal : dLys [33], podczas gdy modele CLP mogą je zaniżać [34]. W niniejszej pracy, szacowane zapotrzebowanie dla ADG w fazie starter nie było zależne od modelu statystycznego (0,68 model EA: 95% maksymalnej odpowiedzi i 0,67 przy użyciu modelu CLP). Wydaje się więc prawdopodobne, że zapotrzebowanie na Wal dla prosiąt wynosi około 0,68, co jest zgodne z wcześniejszymi badaniami [26].
WNIOSKI
U odsadzonych prosiąt, niedobór Wal jest związany z niższymi przyrostami masy ciała i pobraniem paszy. Suplementacja waliny do paszy podstawowej z jej niedoborem łagodzi obniżenie parametrów użytkowych. Optymalny stosunek dWal : dLys dla przyrostu masy ciała w fazie poodsadzeniowej szacuje się na 0,68, co jest zgodne z wcześniejszymi szacunkami.


















