

 Melissa Izabel Hannas
Melissa Izabel Hannas
Universidade Federal de Viçosa
Trzy aminokwasy rozgałęzione (BCAA) – leucyna (Leu), izoleucyna (Ile) i walina (Wal) – odgrywają istotną rolę w żywieniu świń ze względu na swoje funkcje metaboliczne i konieczność ich spożywania. Ich rola w katabolizmie jest wyjątkowa, ponieważ zależą od enzymów obecnych w mięśniach szkieletowych oraz tkankach pozawątrobowych, gdzie uczestniczą w syntezie białek mięśniowych, produkcji energii i regulacji procesów odpornościowych.
Dopiero na początku XIX wieku BCAA zostały uznane za źródła energii dla ssaków. Leu jest przekształcana w α-acetooctan i β-hydroksymaślan, Wal jest przekształcana w glukozę poprzez bursztynian jako produkt pośredni, a Ile może być metabolizowana zarówno do glukozy lub ciał ketonowych u zwierząt (Wu, 2022). W żywieniu świń aminokwasy rozgałęzione (BCAA) powinny być dostarczane wraz z paszą w proporcjach odpowiadających ich optymalnemu stosunkowi aminokwasów, zgodnie z fazą wzrostu i potencjałem zwierzęcia. W warunkach produkcyjnych trzody chlewnej proporcje BCAA w paszy dla świń mogą być trudne do osiągnięcia. Jednym z czynników, na które należy zwrócić uwagę, jest mieszanie różnych źródeł białka, które mogą być niezbilansowane w zależności od stężenia aminokwasów. Rosnąca potrzeba ograniczania zawartości białka ogólnego w paszy prowadzi do niedoborów aminokwasów i/lub występowania antagonizmu między nimi. Ponadto zmiany w idealnym profilu aminokwasowym w określonych warunkach, takich jak problemy immunologiczne i środowiskowe, mogą utrudniać zaspokojenie zapotrzebowania na BCAA. W niniejszym artykule omówiono badania przeprowadzone w ciągu ostatnich 10 lat nad aminokwasami rozgałęzionymi (BCAA) (Leu, Ile i Wal) u świń. Zwrócono w nim uwagę na wpływ powszechnie stosowanych w żywieniu świń relacji BCAA w fazach odchowu, wzrostu i tuczu, reakcje fizjologiczne oraz morfologię krwi, a także na wyzwania wynikające ze stosowania wysokich poziomów Leu, i co najważniejsze, na praktyczne aspekty suplementacji BCAA w żywieniu świń.

Znaczenie aminokwasów rozgałęzionych
Aminokwasy rozgałęzione (BCAA) – leucyna (Leu), izoleucyna (Ile) i walina (Wal) – są niezbędne w żywieniu świń ze względu na ich wyjątkową rolę w syntezie białek mięśniowych, przemianie energetycznej i wpływie na ogólny stan zdrowia. W przeciwieństwie do innych aminokwasów, BCAA są metabolizowane bezpośrednio w mięśniach szkieletowych, Stanowią one około 35% niezbędnych aminokwasów w białkach mięśniowych, co sprawia, że są one szczególnie ważne dla stymulowania wzrostu tkanki beztłuszczowej i poprawy wykorzystania paszy (Wu i in., 2016). Oprócz tego, że są najobficiej występującymi aminokwasami egzogennymi, BCAA uczestniczą w syntezie związków azotowych, regulują metabolizm glukozy i lipidów, biorą udział w syntezie białka, wspierają zdrowie jelit oraz funkcjonowanie układu odpornościowego poprzez szlak sygnałowy PI3K/AKT/mTOR. Poziom i równowaga BCAA są ściśle związane z różnymi parametrami zdrowia i chorób (Nie i in., 2018).
BCAA stanowią około 35% niezbędnych aminokwasów w białkach mięśniowych Tabela 1, a ich wyjątkowy metabolizm w mięśniach szkieletowych ssaków i ptaków sprawia, że są one kluczowe dla wzrostu i zdrowia.
 Tab. 1. Zawartość aminokwasów rozgałęzionych (BCAA) w tkankach trzody chlewnej, %
Tab. 1. Zawartość aminokwasów rozgałęzionych (BCAA) w tkankach trzody chlewnej, %
Katabolizm BCAA obejmuje trzy wspólne etapy: transaminację, oksydacyjną dekarboksylację oraz dehydrogenację acyl-CoA, z których pierwsze są reakcjami o wysokim powinowactwie do tkanek. U ssaków i ptaków poziom enzymu białkowego cytozolowego w wątrobie jest znacznie niższy niż w mięśniach szkieletowych, nerkach, sercu i jelicie cienkim, co wskazuje, że transaminacja w wątrobie jest ograniczona przy normalnych stężeniach fizjologicznych BCAA. BCAA są dostępne dla tkanek pozawątrobowych gdzie uczestniczą w syntezie glutaminianu, glutaminy i alaniny, aktywacji szlaku sygnałowego mTOR oraz stymulacji syntezy białek (Wu, 2022).
Leu w szczególności aktywuje szlak mTOR, stymulując rozwój mięśni, podczas gdy Ile i Wal wspierają równowagę azotową i zapobiegają katabolizmowi mięśni, zapewniając optymalną wydajność wzrostu u świń (Duan i in., 2021). Aktywacja szlaku mTOR przez Leu stymuluje syntezę białek mięśniowych o 20-30% skuteczniej niż inne aminokwasy. Ile odgrywa kluczową rolę w metabolizmie energetycznym i funkcjonowaniu układu odpornościowego, szczególnie w okresie po odsadzeniu. Jako aminokwas glukogeniczny, przyczynia się do 12% produkcji glukozy w wątrobie u prosiąt poddanych głodówce (Bertolo i in., 2008). Biorąc pod uwagę dostępne źródła białka stosowane w paszach dla świń, Wal plasuje się na 3. do 5. miejscu wśród aminokwasów ograniczających.
W ostatnim dziesięcioleciu zainteresowanie badaniami nad BCAA wzrosło ze względu na dążenie do ograniczenia zawartości białka ogólnego w paszach, co prowadzi do zaburzenia równowagi aminokwasowej i/lub niedoborów, a także ze względu na dostępność źródeł białka z nadmiarem leucyny, co powoduje konkurencję i/lub antagonizm w przyswajaniu BCAA, jak również dostępność krystalicznych źródeł aminokwasów do suplementacji. W tej sytuacji przeprowadzono badania, które oszacowały wpływ suplementacji, niedoborów oraz interakcji między BCAA na wydajność oraz reakcje fizjologiczne i metabolomiczne, co pozwoliło określić optymalne proporcje tych aminokwasów w paszy dla świń.
Zalecenia dotyczące BCCA w paszach dla świń
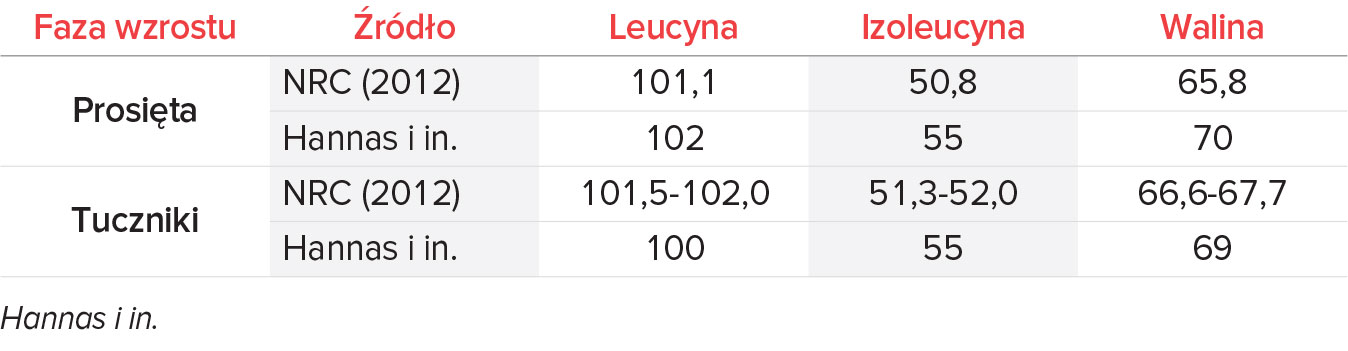
Wysokie zapotrzebowanie prosiąt na BCAA wynika z szybkiego przyrostu masy mięśniowej i dużej zdolności do przyrostu tkanki beztłuszczowej, dodatkowo spotęgowanego przez ich ograniczoną zdolność trawienną. Jako punkt odniesienia przeanalizowano znormalizowany profil aminokwasów BCAA w diecie świń różnych grup wiekowych, który został przytoczony w publikacjach dotyczących żywienia zwierząt, takich jak NRC (2012) oraz Hannas i in. (2024) (Tabela 2). Źródła te stanowią wytyczne do określenia poziomów składników pokarmowych stosowanych w żywieniu świń, jednak powinny być regularnie aktualizowane i uzupełniane o wyniki badań uwzględniające różne grupy zwierząt, różne modele oceny, wyzwania i/lub interakcje. Tylko wtedy możliwe jest precyzyjne żywienie, które pozwala osiągać lepsze wyniki produkcyjne i zdrowotne.
Tab. 2. Zalecenia dotyczące stosunku leucyny, izoleucyny i waliny w odniesieniu do SID Lizyny dla świń
Optymalizacja zawartości BCAA w paszach dla świń
Skupienie się na fazach odchowu i Grower, Finiszer.
Odpowiednia suplementacja waliny w stosunku do lizyny wpływa na wydajność i zdrowie jelit prosiąt po odsadzeniu. Według badań przeprowadzonych przez Xu i wsp. (2018) przyrost masy ciała prosiąt wykazał lepszą reakcję przy poziomie 68% SID Wal : Lys, czyli wartości wyższej niż zalecenia NRC (2012). Lepszym wynikom towarzyszył wzrost wysokości wypustków jelitowych dwunastnicy, jelita czczego i jelita krętego oraz wzrost stosunku wysokości wypustków do głębokości krypt, co wskazuje na większą zdolność wchłaniania i regenerację komórek jelitowych. Jednocześnie zaobserwowano wzrost stężenia Wal i α-ketoisowalerianu w osoczu, przy jednoczesnym spadku stężenia innych aminokwasów, takich jak asparaginian i asparagina. Badania wykazały również, że stosunek SID Wal : Lys zwiększał ekspresję genów enzymów BCATm i BCKDH w mięśniu najdłuższym grzbietu, co świadczy o adaptacji metabolizmu aminokwasów rozgałęzionych. Wyniki te sugerują, że optymalny stosunek SID Wal : Lys maksymalizuje wzrost i sprzyja poprawie struktury jelit oraz adaptacji metabolicznej, podkreślając istotną rolę Val poza syntezą białek. Odkrycia te uzasadniają ponowne rozważenie zaleceń żywieniowych dla prosiąt odsadzonych, biorąc pod uwagę ich kompleksowy wpływ na wydajność i fizjologię trawienia.
Wykorzystując różne modele oceny stosunku aminokwasów, Nørgaard i wsp. (2017) wykazali, że analiza metabolitów w osoczu pozwala zidentyfikować istotne reakcje biologiczne na różne poziomy Ile, Leu i Wal u prosiąt o masie ciała od 8 do 9 kg. Spośród 18 analizowanych aminokwasów w osoczu 11 dla Ile, 16 dla Leu i 3 dla Wal wykazało reakcje liniowe lub kwadratowe. Jednocześnie kluczowe metabolity, takie jak 3-metylo-2-oksowalerian i α-ketoisowalerian, były podatne na zmiany w diecie. Optymalne poziomy oszacowane przez modele metabolomiczne wynosiły 104% SID Leu : Lys, 54 SID Ile : Lys i 68% SID Wal : Lys, co jest zgodne z doniesieniami w literaturze.
Odpowiedni stosunek BCAA, oprócz wpływu na wydajność prosiąt, wydaje się być związany ze zmiennością spożycia paszy, jak stwierdzili Tian i wsp. (2019), gdzie stosowanie paszy ubogiej w aminokwasy rozgałęzione (BCAA) zmniejszało pobór paszy przez prosięta poprzez dwa główne mechanizmy opisane przez autorów: regulację receptorów aminokwasowych T1R1/T1R3 w jelicie oraz aktywację szlaków GCN2/eIF2α w podwzgórzu, a odpowiednia suplementacja BCAA (0,13% L-Ile, 0,09% L – Leu i 0,23% L-Wal) w paszy o niskiej zawartości białka (17,05% CP) zwiększała pobór paszy, co wiązało się ze zmniejszeniem wydzielania cholecystokininy i modulacją szlaków sygnałowych podwzgórza, podczas gdy nadmierna suplementacja BCAA (150% zapotrzebowania) paradoksalnie zmniejszała pobór paszy, co świadczy o znaczeniu prawidłowej proporcji tych aminokwasów w regulacji poboru paszy.
Chociaż stosunki BCAA są określone dla zwierząt w różnych grupach, Veira (2021) podkreślił w swoich eksperymentach, że reakcje na proporcje BCAA u prosiąt wykazywały znaczne różnice w zależności od osobnika, gdzie optymalny stosunek SID Wal : Lys wahał się między 62 a 68% między poszczególnymi osobnikami, ze średnią 67% przy zastosowaniu modelu liniowego i 70% przy zastosowaniu modelu kwadratowego, podczas gdy stosunek SID Ile : Lys wynosił od 52 do 60%, ze średnią odpowiednio 55 i 62% dla modeli liniowego i kwadratowego. Retencja azotu na maksimum plateau różniła się znacznie między zwierzętami (34,78-52,03 g dla Wal i 36,44-56,04 g dla Ile), co podkreśla znaczenie zmienności indywidualnej w precyzyjnym żywieniu. Wyniki pokazują, że zapotrzebowanie na Wal i Ile może się różnić między świniami w tym samym badaniu, z wahaniami do 10% dla Wal i 15% dla Ile, co wskazuje, że różne podejścia statystyczne mogą również wpływać na zalecenia dotyczące składników odżywczych. Podobne wyniki przedstawili Millet i in. (2021), którzy stwierdzili, że idealny stosunek SID Wal : Lys wynosił od 68 do 74% w celu maksymalizacji wydajności prosiąt, przy czym zalecane proporcje różniły się nawet o 6% w zależności od modelu statystycznego.
Tab. 3. Równania regresji do przewidywania wydajności wzrostu tuczników (Cemin i in. 2019)
Badania podkreślają interakcję między aminokwasami rozgałęzionymi (BCAA), a w niektórych sytuacjach nadmiar leucyny może hamować wchłanianie izoleucyny i waliny, co prowadzi do obniżenia wydajności, jeśli proporcje nie są odpowiednio zachowane (Wiltafsky i in., 2009). Antagonizm aminokwasów rozgałęzionych może występować podczas transportu i/lub katabolizmu aminokwasów (Wu, 2022). Suplementacja SID Wal : Lys sprzyjała poprawie przyrostu masy ciała i spożycia paszy u prosiąt, zmniejszając współczynnik konwersji paszy, zgodnie z badaniami Zouaoui i in. (2021). Nadmiar leucyny wywierał znaczny efekt antagonistyczny, zmniejszając przyrost masy ciała i spożycie paszy u świń oraz intensyfikując reakcję na walinę, zwłaszcza w paszach o wysokiej zawartości leucyny. Ten wzorzec sugeruje, że leucyna moduluje przede wszystkim spożycie paszy, a nie jej wydajność. Autorzy podkreślili, że opracowane modele pozwalają dokładnie określić wpływ Leu na zapotrzebowanie na Wal i że kluczowe znaczenie ma uwzględnienie jej poziomu w celu optymalizacji pasz o niskiej zawartości białka, a równowaga między BCAA powinna być dostosowana z uwzględnieniem konkretnych interakcji, przy czym Leu jest głównym regulatorem katabolizmu tych aminokwasów.
Badając złożone interakcje między leucyną, izoleucyną i waliną u prosiąt w fazie odchowu, z wykorzystaniem modelu doświadczalnego obejmującego 15 różnych dawek, w których zmieniano proporcje SID BCAA : lizyna, Humphrey i in. (2023) odkryli, że walina wywierała liniowy i kwadratowy wpływ na dzienny przyrost masy ciała i wykorzystanie paszy, niezależnie od poziomu leucyny i izoleucyny w diecie. Jednak zaobserwowano znaczącą interakcję między Leu i Ile, gdzie jednoczesny wzrost obu aminokwasów pogarszał wyniki, podczas gdy dostosowanie proporcji (wzrost jednego przy zmniejszeniu drugiego) poprawiało przyrost masy ciała i konwersję paszy. Jednym z najbardziej istotnych odkryć była interakcja między Leu i Wal na spożycie paszy; gdy poziomy Wal były poniżej zaleceń NRC (2012), wzrost Leu zmniejszał spożycie paszy. Jednak ten niekorzystny efekt został całkowicie zneutralizowany, gdy podawano odpowiednią ilość waliny lub dawkę wyższą od zalecanej. Sugeruje to, że odpowiednia suplementacja waliny może zrównoważyć negatywny wpływ nadmiaru leucyny na spożycie paszy, pokazując, że optymalną wydajność prosiąt można osiągnąć dzięki różnym kombinacjom aminokwasów rozgałęzionych (BCAA), w zależności od celu produkcji i kosztu składników.
Wzajemne oddziaływanie między BCAA a odpowiedzią immunologiczną świń zostało zbadane przez Rudara i wsp. (2016 i 2018). Oceniając wpływ suplementacji leucyną na retencję azotu u prosiąt przed i po ekspozycji na lipopolisacharyd (LPS), Rudar i wsp. (2016) zaobserwowali, że u zdrowych świń podawanie wysokich dawek leucyny wywoływało reakcję anaboliczną kompensującą zmniejszoną retencję azotu przy średnich poziomach leucyny.
Następnie Rudar i wsp. (2018) wykazali, że stres immunologiczny i suplementacja leucyną znacząco zmieniły metabolizm białek u prosiąt, wywierając wyraźny wpływ na różne tkanki. Stres immunologiczny spowodował gwałtowny wzrost stężenia niezbędnych aminokwasów i mocznika w osoczu w ciągu pierwszych 12 godzin oraz zwiększył tempo syntezy białek w wątrobie, podczas gdy suplementacja leucyną (do 2,72% SID Leu) miała tendencję do zmniejszania tempa syntezy białek w wątrobie i zwiększania jej w mięśniu dwugłowym łydki u zwierząt poddanych stresowi immunologicznemu, co sugeruje redystrybucję aminokwasów do syntezy białek mięśni obwodowych. Ponadto suplementacja leucyną spowodowała liniowy spadek stężenia izoleucyny, waliny, glutaminy i mocznika w osoczu, co wskazuje na wzrost katabolizmu tych aminokwasów, efekt ten został spotęgowany przez stres immunologiczny. Wyniki sugerują, że manipulowanie stosunkiem SID Leu : Lys może być metodą na lepsze wykorzystanie aminokwasów podczas obciążenia immunologicznego, co sprzyja odkładaniu się białka w mięśniach zamiast syntezy w wątrobie, chociaż potrzebne są dalsze badania, żeby ustalić optymalny stosunek, który minimalizuje utratę białka z narządów wewnętrznych i maksymalizuje przyrost mięśni podczas stresu immunologicznego. Wyniki te pokazują, że leucyna może pozytywnie wpływać na metabolizm białek w normalnych warunkach, ale jej suplementacja nie łagodzi efektów katabolicznych wywołanych aktywacją układu odpornościowego.
W badaniach nad świniami w okresie wzrostu (średnia masa ciała od 25 do 45 kg) Gonçalves i in. (2018) ustalili, że stosunek SID Wal : Lys wynosił 73% dla maksymalnego przyrostu masy ciała i 69% dla maksymalnego wykorzystania paszy.
W badaniach z wykorzystaniem metody eliminacji Faria (2019) ustalił, że stosunek BCAA w dietach dla świń o masie odpowiednio 25 i 60 kg wynosił 109 : 57 : 67 i 109 : 56 : 63 dla Leu : Ile : Wal/SID Lys, czyli wartości mieszczące się w granicach podanych w literaturze.
Nadmiar leucyny w paszy znacząco zmienia dostępność innych aminokwasów rozgałęzionych (BCAA) u rosnących świń; u zwierząt karmionych dietą zawierającą dodatkowe 0,43% L-leucyny zaobserwowano wzrost stężenia leucyny w surowicy i jednoczesny spadek poziomu izoleucyny i waliny w fazie wchłaniania i po wchłonięciu (Morales i in., 2018). Autorzy stwierdzili, że nierównowaga została całkowicie odwrócona po uzupełnieniu diety o 0,20% L-Ile i 0,25% L-Wal, co pokazuje, że nadmiar BCAA w połączeniu może przeciwdziałać niekorzystnym skutkom samego nadmiaru leucyny. Wyniki sugerują, że nadmiar Leu ogranicza dostępność Ile i Wal poprzez dwa główne mechanizmy: zmniejszenie wchłaniania jelitowego i zwiększenie tempa degradacji komórkowej tych aminokwasów. Badanie podkreśla, że strategiczna suplementacja Ile i Wal może skorygować te zaburzenia równowagi metabolicznej. Jednak w warunkach kontrolowanego spożycia pokarmu, jak w tym eksperymencie, efekty te nie przekładają się na różnice w wydajności produkcyjnej.
Biorąc pod uwagę wpływ suplementacji BCAA i ich interakcje, Cemin i wsp. (2019) przeprowadzili kompleksową metaanalizę, wykorzystując dane z 25 artykułów opublikowanych w latach 1995–2018, obejmujących łącznie 44 badania i 210 obserwacji, w celu oceny wpływu BCAA i ich interakcji na wydajność rosnących świń. Dzięki modelowaniu statystycznemu naukowcy opracowali równania prognostyczne dotyczące dziennego przyrostu masy ciała, dziennego spożycia paszy i konwersji paszy, biorąc pod uwagę takie zmienne, jak masa ciała, białko ogólne, błonnik i wielokrotne relacje między aminokwasami (wyrażone w przeliczeniu na strawność). Ostateczne równania uwzględniały określone stosunki, takie jak Leu : Lys, Ile : Lys i Wal : Lys, wykazując, że wzrost stosunku Leu : Lys ma negatywny wpływ na dzienny przyrost masy ciała z powodu braku równowagi z innymi aminokwasami rozgałęzionymi (BCAA) i dużymi aminokwasami neutralnymi (LNAA) Tabela 3. Główne wyniki wykazały, że wysoki poziom Leu w paszy pogarsza wydajność poprzez zmniejszenie współczynnika konwersji paszy i spożycia paszy, zwłaszcza w przypadku względnego niedoboru innych aminokwasów rozgałęzionych, takich jak Wal i Ile, lub aminokwasów neutralnych, takich jak tryptofan (Trp). Badanie wykazało, że strategiczne uzupełnianie Wal, Ile i Trp może złagodzić te niekorzystne skutki poprzez zrównoważenie proporcji między aminokwasami. Opracowany model pozwala przewidywać reakcje produkcyjne w oparciu o relacje między aminokwasami, oferując cenne narzędzia do optymalizacji diety i potwierdzając, że strategie żywieniowe powinny uwzględniać nie tylko bezwzględne poziomy BCAA, ale przede wszystkim ich względne proporcje, co ma praktyczne znaczenie dla producentów trzody chlewnej w celu maksymalizacji wydajności i wyników.
Tab. 4.
Badając wpływ nadmiaru leucyny na metabolizm BCAA u rosnących świń, Kwon i wsp. (2019) wykazali, że dieta o wysokiej zawartości leucyny (100–300% zapotrzebowania) zmniejszała spożycie paszy, retencję azotu i syntezę serotoniny w podwzgórzu, jednocześnie zwiększając stężenie mocznika w osoczu. Nadmiar leucyny powoduje zaburzenia równowagi metabolicznej, utrudniając wykorzystanie izoleucyny i waliny do syntezy białek. Zaobserwowano znaczące zmiany w rozmieszczeniu BCAA w tkankach, gdzie stężenie BCAA w wątrobie wzrosło, a stężenie w mięśniach spadło. Ponadto nastąpiło zmniejszenie poziomu α-ketokwasów pochodzących z waliny i izoleucyny oraz wzrost poziomu pochodnych leucyny, co sugeruje, że nadmiar leucyny przyspiesza degradację izoleucyny i waliny. Wzrosła również ekspresja enzymów katabolicznych (transaminaza BCAA i BCKDH) w mięśniach, co potwierdza hipotezę o większym katabolizmie tych aminokwasów. W badaniu zwrócono również uwagę na wpływ neuroendokrynologiczny: liniowy spadek serotoniny w podwzgórzu i osoczu sugeruje, że nadmiar leucyny zaburza wchłanianie tryptofanu w mózgu, wpływając na regulację apetytu. To zmniejszenie poziomu serotoniny może wyjaśniać spadek dobrowolnego spożycia pokarmu, co w połączeniu z mniejszą dostępnością Val i Ile do syntezy białek przyczyniło się do pogorszenia wyników. Badania wykazały, że nadmiar leucyny wywołuje antagonizm między aminokwasami rozgałęzionymi (BCAA), zmniejszając skuteczność białek i wzrost poprzez trzy główne ścieżki: zwiększony rozkład Wal i Ile, dysregulację sygnalizacji serotoninergicznej w mózgu oraz zmniejszone spożycie pokarmu. Wyniki te potwierdzają potrzebę starannego zrównoważenia stosunku leucyny, waliny i izoleucyny w recepturach paszowych, aby uniknąć zaburzeń metabolicznych i produkcyjnych.
Podsumowanie wyników badań dotyczących BCAA
Dane przedstawione w tabeli 4 podsumowują najważniejsze wyniki badań, podkreślając interakcje między BCAA, optymalne wymagania i wpływ na wydajność zwierząt. Wyniki te stanowią cenne wsparcie przy opracowywaniu bardziej precyzyjnych diet, uwzględniających średnie wartości dla stada i indywidualne różnice w wykorzystaniu aminokwasów. Wyniki podkreślają złożoność metaboliczną BCAA i znaczenie uwzględnienia ich interakcji w formułowaniu pasz.
Wnioski
Badania przeprowadzone w ostatnich latach sugerują, że stosunki BCAA mogą różnić się od standardowych wartości białka, biorąc pod uwagę współczesne genotypy charakteryzujące się wysokim odkładaniem chudego mięsa oraz interakcje z BCAA. W niektórych sytuacjach wyższe lub niższe stężenia BCAA mogą prowadzić do obniżenia wydajności, jeśli nie są odpowiednio zrównoważone. Pasze niskobiałkowe uzupełnione krystalicznymi aminokwasami są coraz częściej stosowane w celu zmniejszenia wydalania azotu; pasze te mogą sprzyjać niedoborom BCAA, jeśli nie są odpowiednio skomponowane. Wal i Ile stają się ograniczające w mieszankach niskobiałkowych, a nieodpowiednia suplementacja może zmniejszyć spożycie paszy i spowolnić wzrost. Podobnie, nadmiar Leu w paszach niskobiałkowych może powodować zaburzenia równowagi BCAA, pogłębiając niedobory Ile i Wal. W praktyce źródła białka w diecie różnią się zawartością BCAA; diety oparte na mączce kukurydzianej i sojowej zazwyczaj dostarczają wystarczającą ilość Leu, ale mogą nie zapewniać odpowiedniej ilości Wal i Ile w stosunku do Lys. Alternatywne białka, takie jak pszenica i śruta rzepakowa, mają różne profile BCAA, co wymaga dostosowań. Nadmiar Leu zmniejsza dostępność Ile, Wal i Trp, wpływając na wzrost i spożycie paszy. Strategiczna suplementacja Wal, Ile lub Trp może skorygować zaburzenia równowagi, zwłaszcza w dietach zawierających kukurydzę/DDGS, gdzie występuje nadmiar Leu.





